Record Number
3008
PROSEA Handbook Number
11: Auxiliary plants
Taxon
Erythrina poeppigiana (Walpers) O.F. Cook
Protologue
Bull. U.S. Dept. Agric. Div. Bot. 25: 57 (1901).
Family
LEGUMINOSAE - PAPILIONOIDEAE
Chromosome Numbers
2n = 42
Synonyms
Erythrina micropteryx Poeppig ex Walpers (1850), Micropteryx poeppigiana Walpers (1850).
Vernacular Names
Mountain immortelle, coral tree (En). Bois immortelle (Fr). Poró gigante, bucayo gigante (Sp). Indonesia: dadap. Similar vernacular names often refer to other Erythrina spp. as well.
Origin and Geographic Distribution
Erythrina poeppigiana occurs naturally in South America, from Venezuela and Panama in the north, throughout the Andean foothills of Colombia, Ecuador, Peru and Bolivia to the western parts of the Amazon basin in the south. It is now extensively planted and naturalized in Central America and the Caribbean. It has been introduced into the humid tropics of the Old World, including South-East Asia.
Uses
Erythrina poeppigiana is one of the most commonly planted shade trees in cocoa, coffee and pepper plantations in Central America, often planted in combination with the agroforestry tree Cordia alliodora (Ruiz & Pavon) Oken. It is valued for its high production of green manure and mulch, the ease with which shade can be adjusted to the requirements of the main crop and its ability to tolerate regular coppicing for many years. It is used occasionally as a shade tree in Indonesia.
It is used less frequently in live fences and as a shade and forage tree in pastures, e.g. in association with Cynodon plectostachyus (K. Schum.) Pilger, Cynodon nlemfuensis Vanderyst and Pennisetum purpureum Schumach. Leaves are cut for fodder for cattle and goats. Pigs are reported to suffer from hair loss when fed with Erythrina poeppigiana loppings.
Seeds and leaves are reportedly used medicinally in the countries of origin. Seeds also yield a fish poison. Like several other Erythrina spp., trees are often grown as ornamentals for their bright orange-red flowers. In Colombia, the flowers are eaten in salads and soups.
The wood is of very limited value, even making only poor quality firewood. It is used occasionally for poles and posts, sometimes for vegetable crates, pulp or particle board. Lopped branches have no value as wood.
It is used less frequently in live fences and as a shade and forage tree in pastures, e.g. in association with Cynodon plectostachyus (K. Schum.) Pilger, Cynodon nlemfuensis Vanderyst and Pennisetum purpureum Schumach. Leaves are cut for fodder for cattle and goats. Pigs are reported to suffer from hair loss when fed with Erythrina poeppigiana loppings.
Seeds and leaves are reportedly used medicinally in the countries of origin. Seeds also yield a fish poison. Like several other Erythrina spp., trees are often grown as ornamentals for their bright orange-red flowers. In Colombia, the flowers are eaten in salads and soups.
The wood is of very limited value, even making only poor quality firewood. It is used occasionally for poles and posts, sometimes for vegetable crates, pulp or particle board. Lopped branches have no value as wood.
Properties
The edible biomass contains per 100 g dry matter (22—)27—34 g crude protein with an in vitro digestibility of 50—80%. Mineral content of prunings per 100 g dry matter is: leaves: N 3.1 g, P 0.24 g, K 1.3 g, Ca 1.6 g, Mg 0.5 g; branches: N 1.2 g, P 0.15 g, K 1.3 g, Ca 1.25 g, Mg 0.4 g. Fallen leaves contain per 100 g dry matter: N 2.2—2.6 g, P 0.14—0.15 g, K 0.5—0.6 g, Ca 1.9—2.2 g, Mg 0.5—0.7 g. Branches in natural litter contain per 100 g dry matter: N 1.3 g, P 0.1 g, K 0.7 g, Ca 2.0 g, Mg 0.7 g. Mineral content is influenced by the pruning regime. Frequent pruning increases the proportion of leaves and the N content of the loppings.
Like other Erythrina spp., Erythrina poeppigiana contains curare-like alkaloids with a muscle relaxant or paralysing action and uncommon, non-protein amino acids assumed to have insecticidal properties. The alkaloids have been subject of extensive pharmaceutical tests. Species can be identified by the alkaloid profile of the seeds. Erythrina poeppigiana contains erysotrine and its derivatives erythratidine and erythroidine. These alkaloids are not poisonous to ruminants in the quantities present in normal rations.
The wood is characterized by abundant thin-walled, axial and radial parenchyma. It is soft, light in weight (specific gravity 250 kg/m3), whitish to yellowish. Heartwood can not be distinguished from sapwood. Large vessels, the prominent rays and parenchyma bands are easily visible using a hand lens.
The seed weight of 100 seeds is 15—30 g.
Like other Erythrina spp., Erythrina poeppigiana contains curare-like alkaloids with a muscle relaxant or paralysing action and uncommon, non-protein amino acids assumed to have insecticidal properties. The alkaloids have been subject of extensive pharmaceutical tests. Species can be identified by the alkaloid profile of the seeds. Erythrina poeppigiana contains erysotrine and its derivatives erythratidine and erythroidine. These alkaloids are not poisonous to ruminants in the quantities present in normal rations.
The wood is characterized by abundant thin-walled, axial and radial parenchyma. It is soft, light in weight (specific gravity 250 kg/m3), whitish to yellowish. Heartwood can not be distinguished from sapwood. Large vessels, the prominent rays and parenchyma bands are easily visible using a hand lens.
The seed weight of 100 seeds is 15—30 g.
Description
A sometimes multi-stemmed, deciduous, often spiny tree with spreading crown, up to 25 m tall and 1.2 m in trunk diameter; when cultivated it is generally kept small, by cutting the stem to 2—2.5 m. Bark greenish-brown to grey-brown, nearly smooth or slightly furrowed, warty or spiny. Twigs stout, spiny, light green and puberulous when young, becoming greenish-grey, with raised leaf scars. Leaves alternate, trifoliolate, thin-chartaceous, often scabrous beneath, glabrescent; petiole 10—40 cm long; rachis up to 30 cm long, with cup-like stipellar, nectar-producing glands at the base of lateral leaflets; petiolules up to 1.5 cm long; leaflets ovate to rhombic, terminal one 8—30 cm x 5—30 cm. Inflorescence an axillary raceme, borne horizontally at distal end of shoots, densely and finely tomentellous; peduncle 4—8 cm long; rachis 7—40 cm long; pedicel 0.5—1.2 cm long, very finely tomentellous or puberulent; bracts and bracteoles ovate, up to 1.3 mm x 0.8 mm; calyx campanulate, 5—10 mm long and wide, orange to reddish at the top, entire but with 2 mm long spur on keel side; standard elliptical, 3—5 cm x 1.5—2.5 cm, bright orange, erecto-patent or slightly recurved, claw 1.5 mm long; wings spatulate to obovate, 7—14 mm x 3—6 mm; keel falcate, 3—5 cm x 0.5—1 cm; stamens 10, 1 free, 9 tubular connate at base, 3—5 cm long, separate for 0.5—1 cm; pistil up to 5 cm long, with linear, puberulent ovary and filiform style. Pod 13—25 cm x 1—1.5 cm, chartaceous, not constricted between seeds, stipe 3—4 cm long, beak 4—8 mm. Seed oblongoid to ellipsoid to slightly reniform, 10—17 mm x 5—7 mm, glossy-brown, without markings.
Image
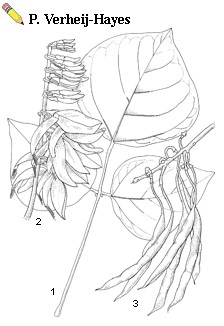 | Erythrina poeppigiana (Walpers) O.F. Cook - 1, leaf; 2, inflorescence; 3, infructescence |
Growth and Development
Mature seed germinates readily. The germination rate is 60—90%, decreasing with storage time. Under favourable conditions, seedlings attain 60—85 cm height and a basal diameter of 2—3 cm in 4 months. In 6 months, trees can attain a height of 4—5 m and develop a crown of 3—4 m diameter with up to 15 branches.
The growth of the tree follows the architectural model of Attim. The main and lateral shoots display indeterminate growth. Most mature trees shed their leaves early in the dry season. Leaf abscission is generally followed by flowering. Flowers open acropetally on newly formed lateral inflorescences at the distal ends of shoots. Fruits develop on leafless trees, maturing in about 2 months, and new shoots emerge during or after fruit maturation. However, this development may be asynchronous within the crown of large trees; on upper branches leaves may abscise and flowers may develop, while lower branches retain their leaves. Trees in some areas shed their leaves again at the end of the wet season, but this leaf fall is not followed by a flush of flowers. Where a dry season is absent, trees are never bare and flower when in leaf. Near the equator trees may flower twice a year. The processes of leaf and flower development seem to be controlled by internal moisture conditions. The increased availability of moisture immediately after leaf fall has been suggested as a stimulus for flowering.
Erythrina poeppigiana is capable of atmospheric nitrogen fixation by nodulation with Bradyrhizobium. Successful inoculation has been obtained with strain CIAT 71. Nodulation starts early and plantlets may contain up to 80 nodules 3 months after sowing. Nodules are 1.5—10 mm in diameter, spherical and clustered on the central root system at the point of emergence of lateral roots. They occur only in the top 10—12 cm of the soil, mainly in the area under the crown of the tree. The biomass of the root nodules varies from 80—250 mg dry matter per dm3 soil and is largest close to the stem.
Pollarding affects nitrogen fixation of the trees. Some of the root nodules and part of the root system disintegrate after the tree has been pruned. New roots and nodules form when the development of new foliage is well under way.
The growth of the tree follows the architectural model of Attim. The main and lateral shoots display indeterminate growth. Most mature trees shed their leaves early in the dry season. Leaf abscission is generally followed by flowering. Flowers open acropetally on newly formed lateral inflorescences at the distal ends of shoots. Fruits develop on leafless trees, maturing in about 2 months, and new shoots emerge during or after fruit maturation. However, this development may be asynchronous within the crown of large trees; on upper branches leaves may abscise and flowers may develop, while lower branches retain their leaves. Trees in some areas shed their leaves again at the end of the wet season, but this leaf fall is not followed by a flush of flowers. Where a dry season is absent, trees are never bare and flower when in leaf. Near the equator trees may flower twice a year. The processes of leaf and flower development seem to be controlled by internal moisture conditions. The increased availability of moisture immediately after leaf fall has been suggested as a stimulus for flowering.
Erythrina poeppigiana is capable of atmospheric nitrogen fixation by nodulation with Bradyrhizobium. Successful inoculation has been obtained with strain CIAT 71. Nodulation starts early and plantlets may contain up to 80 nodules 3 months after sowing. Nodules are 1.5—10 mm in diameter, spherical and clustered on the central root system at the point of emergence of lateral roots. They occur only in the top 10—12 cm of the soil, mainly in the area under the crown of the tree. The biomass of the root nodules varies from 80—250 mg dry matter per dm3 soil and is largest close to the stem.
Pollarding affects nitrogen fixation of the trees. Some of the root nodules and part of the root system disintegrate after the tree has been pruned. New roots and nodules form when the development of new foliage is well under way.
Other Botanical Information
The species of Erythrina L. can, as far as is known, all be intercrossed to produce fertile hybrids.
Erythrina poeppigiana is pollinated by many non-specialized perching or sparrow-like birds. The amount of nectar in the flowers may reach 50 µg per flower and is so great that insects would visit too few flowers to be effective distributors of pollen.
Erythrina poeppigiana is pollinated by many non-specialized perching or sparrow-like birds. The amount of nectar in the flowers may reach 50 µg per flower and is so great that insects would visit too few flowers to be effective distributors of pollen.
Ecology
In cultivation, Erythrina poeppigiana can adapt to a wide range of conditions. It is found from (0—)500—1500(—2000) m above sea level in the tropics with annual rainfall ranging from 1200—3000 mm, with a period of up to 6 months of reduced rainfall. Average annual temperatures may vary from 18—28°C. Above 2000 m altitude, trees become blanketed with epiphytes and stunted, but may survive up to 2400 m.
It will grow on a wide range of soils, from heavy clay to medium loam, ranging from acid to alkaline, but very acid soils are not tolerated.
Trees are resistant to fire, including controlled burning.
It will grow on a wide range of soils, from heavy clay to medium loam, ranging from acid to alkaline, but very acid soils are not tolerated.
Trees are resistant to fire, including controlled burning.
Propagation and planting
Erythrina poeppigiana is sometimes propagated by seed, suckers or air layering, but farmers usually propagate it by large stem cuttings, 2—2.5 m long and 8—12 cm in diameter, readily obtainable from 2-year-old branches. Sprouting starts within a month after planting, and in 4—6 months the new trees start shading coffee seedlings. Rooting success varies from 70—90%. Methods of in vitro propagation are being developed.
Husbandry
Established shade trees in coffee or cocoa plantations are normally completely or partially pollarded once or twice a year. Timing and intensity of pruning can be adjusted to prevailing conditions and the requirements of the associated crop. In coffee plantations, trees are usually pruned prior to flowering and again before ripening of the crop, or at the beginning of the long and the short rainy seasons. Additional pruning is sometimes carried out during prolonged periods of extremely cloudy weather. To maintain a low shade canopy the calloused trunk tip is sawn off every 5—6 years. Trees tolerate pollarding well and can be treated this way for many years.
Erythrina poeppigiana produces large quantities of prunings and litter. Planted at a density of 280 trees per ha in a coffee plantation under experimental conditions in Costa Rica, pollarding once a year produced 18.5 t dry matter per ha of prunings and 4.3 t dry matter per ha of natural litter. Of the prunings 3.2 t were leaves, 15.2 t were branches. When pollarded twice a year, annual dry matter production fell to 11.8 t/ha of prunings and 1.9 t/ha of natural litter. Three prunings per year resulted in an annual dry matter production of 7.9 t/ha of prunings, consisting of 4.3 t of leaves and 3.5 t of branches. No natural litter fall occurred under the latter pruning system, as the life span of leaves exceeds 4 months. Prolonging the pruning interval thus results in a greatly increased production of branch wood and a gradual decrease in the production of leaves. Leaf production of the associated coffee crop is also larger than in coffee grown without shade. In cocoa plantations with a similar density of Erythrina poeppigiana comparable amounts of prunings and litter are found.
The amounts of nutrients recycled in the prunings are considerable and mostly match fertilizer recommendations for intensively managed coffee. In the pollarding experiment mentioned, annual pollarding resulted in contributions per ha of 330 kg N, 32 kg P, 156 kg K, 319 kg Ca, 86 kg Mg. When pollarding three times per year annual contributions per ha were: 173 kg N, 14 kg P, 119 kg K, 94 kg Ca, 27 kg Mg. Amounts for pollarding twice a year were intermediate. Although considerable amounts of nutrients are recycled, large quantities are immobilized in the stems as well. The total amount of available P in the soil and the litter layer may actually diminish under Erythrina poeppigiana. However, when trees are pollarded 2 or 3 times per year, most of the stored nutrients are recycled. It was found that most of the nitrogen recycled was taken up from the soil. N balance studies indicate that up to 60 kg atmospheric N per ha may be assimilated annually, even when the crop receives mineral N fertilizer. This is comparable to several other woody legumes, but less than the quantities reached by Leucaena leucocephala (Lamk) de Wit.
There is little information on the durability of the mulch layer. Studies indicate that 50% of the organic matter decomposes within 1 month and 75% in 6 months. The decomposition of organic material may cause the pH to decline in certain soils, which may result in increased leaching of K, Ca and Mg.
The high production of organic matter in prunings results in an increase in soil organic matter. In a ten-year experiment with Erythrina poeppigiana and cocoa, the soil organic matter increased from a relatively high initial amount of 200 t/ha to 240 t/ha. This contributes to very low levels of leaching of minerals, comparable to those in natural or planted forests.
Neither the intensity nor the effects of the shade of Erythrina poeppigiana have ever been measured systematically and directly.
In long-term trials with maize (Zea mays L.), grown in the rainy season, followed by a dry season crop of common bean (Phaseolus vulgaris L.), both crops showed considerable yield increases in response to an annual mulch of 20 t/ha of Erythrina poeppigiana prunings. The N utilization by the crops was somewhat less efficient than with mineral N fertilizer. The cut-and-carry mulch system was superior to an alley-cropping system with the same crops. The yield advantages were smaller in the alley-cropping system. An economic analysis of the systems showed the cut-and-carry system to be more profitable in spite of the high labour requirements. Cassava (Manihot esculenta Crantz) yields could not be maintained, either in the mulch system or in the alley-cropping system.
Erythrina poeppigiana has been tested as shade and forage tree with a number of grasses, e.g. king grass (a hybrid of Pennisetum purpureum and P. glaucum (L.) R. Br.) and Cynodon nlemfuensis. Yields of the grasses were not reduced and sometimes even increased under Erythrina poeppigiana. The protein content increased. However, after a few years, yields started to decline because of the large loss of nutrients caused by removing cut grass and Erythrina poeppigiana loppings. Leaves of Erythrina poeppigiana are readily accepted by livestock and may increase the amount of feed ingested and milk produced.
Erythrina poeppigiana produces large quantities of prunings and litter. Planted at a density of 280 trees per ha in a coffee plantation under experimental conditions in Costa Rica, pollarding once a year produced 18.5 t dry matter per ha of prunings and 4.3 t dry matter per ha of natural litter. Of the prunings 3.2 t were leaves, 15.2 t were branches. When pollarded twice a year, annual dry matter production fell to 11.8 t/ha of prunings and 1.9 t/ha of natural litter. Three prunings per year resulted in an annual dry matter production of 7.9 t/ha of prunings, consisting of 4.3 t of leaves and 3.5 t of branches. No natural litter fall occurred under the latter pruning system, as the life span of leaves exceeds 4 months. Prolonging the pruning interval thus results in a greatly increased production of branch wood and a gradual decrease in the production of leaves. Leaf production of the associated coffee crop is also larger than in coffee grown without shade. In cocoa plantations with a similar density of Erythrina poeppigiana comparable amounts of prunings and litter are found.
The amounts of nutrients recycled in the prunings are considerable and mostly match fertilizer recommendations for intensively managed coffee. In the pollarding experiment mentioned, annual pollarding resulted in contributions per ha of 330 kg N, 32 kg P, 156 kg K, 319 kg Ca, 86 kg Mg. When pollarding three times per year annual contributions per ha were: 173 kg N, 14 kg P, 119 kg K, 94 kg Ca, 27 kg Mg. Amounts for pollarding twice a year were intermediate. Although considerable amounts of nutrients are recycled, large quantities are immobilized in the stems as well. The total amount of available P in the soil and the litter layer may actually diminish under Erythrina poeppigiana. However, when trees are pollarded 2 or 3 times per year, most of the stored nutrients are recycled. It was found that most of the nitrogen recycled was taken up from the soil. N balance studies indicate that up to 60 kg atmospheric N per ha may be assimilated annually, even when the crop receives mineral N fertilizer. This is comparable to several other woody legumes, but less than the quantities reached by Leucaena leucocephala (Lamk) de Wit.
There is little information on the durability of the mulch layer. Studies indicate that 50% of the organic matter decomposes within 1 month and 75% in 6 months. The decomposition of organic material may cause the pH to decline in certain soils, which may result in increased leaching of K, Ca and Mg.
The high production of organic matter in prunings results in an increase in soil organic matter. In a ten-year experiment with Erythrina poeppigiana and cocoa, the soil organic matter increased from a relatively high initial amount of 200 t/ha to 240 t/ha. This contributes to very low levels of leaching of minerals, comparable to those in natural or planted forests.
Neither the intensity nor the effects of the shade of Erythrina poeppigiana have ever been measured systematically and directly.
In long-term trials with maize (Zea mays L.), grown in the rainy season, followed by a dry season crop of common bean (Phaseolus vulgaris L.), both crops showed considerable yield increases in response to an annual mulch of 20 t/ha of Erythrina poeppigiana prunings. The N utilization by the crops was somewhat less efficient than with mineral N fertilizer. The cut-and-carry mulch system was superior to an alley-cropping system with the same crops. The yield advantages were smaller in the alley-cropping system. An economic analysis of the systems showed the cut-and-carry system to be more profitable in spite of the high labour requirements. Cassava (Manihot esculenta Crantz) yields could not be maintained, either in the mulch system or in the alley-cropping system.
Erythrina poeppigiana has been tested as shade and forage tree with a number of grasses, e.g. king grass (a hybrid of Pennisetum purpureum and P. glaucum (L.) R. Br.) and Cynodon nlemfuensis. Yields of the grasses were not reduced and sometimes even increased under Erythrina poeppigiana. The protein content increased. However, after a few years, yields started to decline because of the large loss of nutrients caused by removing cut grass and Erythrina poeppigiana loppings. Leaves of Erythrina poeppigiana are readily accepted by livestock and may increase the amount of feed ingested and milk produced.
Diseases and Pests
Erythrina poeppigiana is not seriously affected by diseases or pests. It is termite resistant even when regularly pollarded.
Genetic Resources
The Nitrogen Fixing Tree project maintains a collection of 28 species and about 75 accessions of species commonly used in Costa Rica at the Centro Agronómico Tropical de Investigación y Enseñanza (CATIE), Turrialba. Another large collection of species of Erythrina is maintained in the Waimea Arboretum, Haleiwa, Hawaii.
Breeding
At CATIE, Turrialba, selections have been made on the basis of desirable characteristics such as absence of spines, branching habit, capacity to retain leaves during the dry season and value as forage crop. Selection trials are still in progress. As the selections used in Central America seem to be derived from a very limited number of introductions, there is ample scope for including genetic materials from the regions of origin.
Prospects
Its fast growth, ability to produce large amounts of biomass and to fix atmospheric nitrogen, ease of propagation from cuttings, excellent response to pruning and high content of crude protein mean that Erythrina poeppigiana has excellent potential for various agroforestry practices not only in Central America but throughout the humid tropics. It plays a central role in agroforestry and alley-cropping research work at CATIE, Turrialba, Costa Rica. Besides its role as shade tree, it fits in well in alley cropping with annual crops and pasture grasses. Its productivity and the good digestibility of its protein make it a promising forage crop. The low value of the wood is compensated by other qualities.
Literature
Beer, J., 1988. Litter production and nutrient cycling in coffee (Coffea arabica) or cacao (Theobroma cacao) plantations with shade trees. Agroforestry Systems 7: 103-114.
Borchert, R., 1980. Phenology and ecophysiology of tropical trees: Erythrina poeppigiana O.F. Cook. Ecology 61: 1065-1074.
Fassbender, H.W., Beer, J., Heuveldop, J., Imbach, A., Enriquez, G. & Bonneman, A., 1991. Ten year balances of organic matter and nutrients in agroforestry systems at CATIE, Costa Rica. Forest Ecology and Management 45: 173-183.
Krukoff, B.A., 1939. The American species of Erythrina. Brittonia 3: 235-238.
Pezo, D., Kass, M., Benavides, J., Romero, F. & Chaves, C., 1990. Potential of legume tree fodders as animal feed in Central America. In: Devendra, C. (Editor): Shrubs and tree fodders for farm animals. Proceedings of a workshop in Denpasar, Indonesia, 24-29 July, 1989. International Development Research Center of Canada (IDRC), Ottawa, Canada. pp. 163-175.
Ramírez, C., Sanchez, G., Kass, D., Viquez, E., Sanchez, N., Vasquez, N. & Ramirez, G., 1989. Advances in Erythrina research at CATIE. In: Werner, D. & Müller, P. (Editors): Fast growing trees and nitrogen fixing trees. Fisher Verlag, Stuttgart, Germany. pp. 96-105.
Russo, R.O., 1990. Erythrina: a versatile genus for agroforestry systems in the tropics. Journal of Sustainable Agriculture 1: 89-109.
Russo, R.O. & Budowski, G., 1986. Effect of pollarding frequency on biomass of Erythrina poeppigiana as a coffee shade tree. Agroforestry Systems 4: 145-162.
Borchert, R., 1980. Phenology and ecophysiology of tropical trees: Erythrina poeppigiana O.F. Cook. Ecology 61: 1065-1074.
Fassbender, H.W., Beer, J., Heuveldop, J., Imbach, A., Enriquez, G. & Bonneman, A., 1991. Ten year balances of organic matter and nutrients in agroforestry systems at CATIE, Costa Rica. Forest Ecology and Management 45: 173-183.
Krukoff, B.A., 1939. The American species of Erythrina. Brittonia 3: 235-238.
Pezo, D., Kass, M., Benavides, J., Romero, F. & Chaves, C., 1990. Potential of legume tree fodders as animal feed in Central America. In: Devendra, C. (Editor): Shrubs and tree fodders for farm animals. Proceedings of a workshop in Denpasar, Indonesia, 24-29 July, 1989. International Development Research Center of Canada (IDRC), Ottawa, Canada. pp. 163-175.
Ramírez, C., Sanchez, G., Kass, D., Viquez, E., Sanchez, N., Vasquez, N. & Ramirez, G., 1989. Advances in Erythrina research at CATIE. In: Werner, D. & Müller, P. (Editors): Fast growing trees and nitrogen fixing trees. Fisher Verlag, Stuttgart, Germany. pp. 96-105.
Russo, R.O., 1990. Erythrina: a versatile genus for agroforestry systems in the tropics. Journal of Sustainable Agriculture 1: 89-109.
Russo, R.O. & Budowski, G., 1986. Effect of pollarding frequency on biomass of Erythrina poeppigiana as a coffee shade tree. Agroforestry Systems 4: 145-162.
Author(s)
L.P.A. Oyen
Correct Citation of this Article
Oyen, L.P.A., 1997. Erythrina poeppigiana (Walpers) O.F. Cook. In: Faridah Hanum, I & van der Maesen, L.J.G. (Editors): Plant Resources of South-East Asia No 11: Auxiliary plants. PROSEA Foundation, Bogor, Indonesia. Database record: prota4u.org/prosea

All texts are licensed under a Creative Commons Attribution-Noncommercial-Share Alike 3.0 Netherlands License
This license does not include the illustrations (Maps,drawings,pictures); these remain all under copyright.