Record Number
3236
PROSEA Handbook Number
9: Plants yielding non-seed carbohydrates
Taxon
Metroxylon sagu Rottboell
Protologue
Nye Saml. K. Danske Vidensk. Selsk. Skrift. 2: 527—528 (1783).
Family
PALMAE
Chromosome Numbers
2n = 26
Synonyms
Metroxylon rumphii Mart. (1845) with many varieties, designated by Beccari (1918); Metroxylon squarrosum Becc. (1918) with many varieties.
Vernacular Names
True sago palm, sago palm (En). Sagoutier (Fr). Indonesia: pohon sagu (general), pohon rumbia (general), kirai (Sundanese), lapia (Ambonese). Malaysia: rumbia (general), balau (Melanau, Sarawak). Papua New Guinea: sak-sak (Pidgin). Philippines: lumbiya. Burma (Myanmar): tha-gu-bin. Cambodia: chraè saku:. Laos: sa:kh'u: tônz. Thailand: sakhu. Vietnam: sagu.
Note: In the Indonesian-Malay language region, the word 'sagu' denotes the edible starch from the pith of any palm and each of these palms may be called 'pohon sagu' (sago tree). 'Pohon rumbia' designates one of them, namely the 'true' sago palm dealt with here, but the name is not commonly used.
Note: In the Indonesian-Malay language region, the word 'sagu' denotes the edible starch from the pith of any palm and each of these palms may be called 'pohon sagu' (sago tree). 'Pohon rumbia' designates one of them, namely the 'true' sago palm dealt with here, but the name is not commonly used.
Origin and Geographic Distribution
The sago palm probably originates from New Guinea and the Moluccas but has only recently been dispersed for research beyond South-East Asia and the nearby Pacific islands. In Indonesia, the palm is now found in parts of Sulawesi, Kalimantan, Sumatra and West Java, as well as on many smaller islands with a non-seasonal climate, notably the Riau Islands, Nias and the Mentawai Islands. In Malaysia, the palm grows in Sabah, Sarawak and on the Peninsula. Some are found in Brunei and in the Philippines (Mindanao). There are large areas of sago palm in Papua New Guinea. There is also a small area in southern Thailand. Sago palm is found at least as far east as the Solomon Islands and probably the Santa Cruz Islands (species have not been identified with certainty).
The world's largest contiguous sago palm swamps and forests are found in New Guinea, totalling a roughly estimated 5—6 million ha, with 4—5 million ha on the Indonesian part of the island.
The world's largest contiguous sago palm swamps and forests are found in New Guinea, totalling a roughly estimated 5—6 million ha, with 4—5 million ha on the Indonesian part of the island.
Uses
The starch stored in the trunk is a staple food, notably in New Guinea. Usually, wet starch is boiled, fried or roasted, alone or mixed with other foodstuffs, resulting in products of different keeping quality. In Indonesia and Malaysia, the starch is used industrially in the manufacture of cakes and cookies, noodles and kerupuk (crisps), and in the United States for custard powders. Non-food uses include sizing pastes for paper and textiles, and extender in adhesive for plywood. It is a very suitable raw material for further industrial processing, e.g. into high-fructose syrup and ethanol.
The palm has many secondary uses. Whole young trunks, pith and pith refuse are given to animals. The 'bark' of the trunk is used as timber or as fuel. Walls, ceilings and fences can be constructed from the petioles ('gaba-gaba'); the fibrous outer layer of the petioles is used for cordage and to weave mats. The leaflets produce one of the best ataps (roof thatch) available, the main use of the palm in West Java. Young leaflets are made into baskets for the transport and storage of fresh (wet) starch. The growing point of the palm with its surrounding tissues may be eaten raw or cooked (palm cabbage).
The larvae of insects feeding on the pith of the trunk, notably weevils of the genus Rhynchophorus, are eaten raw, boiled or roasted in most places where sago palm is a staple. A mushroom (Volvariella volvacea Fries) which grows on pith refuse is relished in the Moluccas.
The palm has many secondary uses. Whole young trunks, pith and pith refuse are given to animals. The 'bark' of the trunk is used as timber or as fuel. Walls, ceilings and fences can be constructed from the petioles ('gaba-gaba'); the fibrous outer layer of the petioles is used for cordage and to weave mats. The leaflets produce one of the best ataps (roof thatch) available, the main use of the palm in West Java. Young leaflets are made into baskets for the transport and storage of fresh (wet) starch. The growing point of the palm with its surrounding tissues may be eaten raw or cooked (palm cabbage).
The larvae of insects feeding on the pith of the trunk, notably weevils of the genus Rhynchophorus, are eaten raw, boiled or roasted in most places where sago palm is a staple. A mushroom (Volvariella volvacea Fries) which grows on pith refuse is relished in the Moluccas.
Production and International Trade
Of the total sago palm area of 5—6 million ha, only an estimated 210 000 ha is planted. Planted areas are estimated to be 130 000 ha in Indonesia, 40 000 ha in Malaysia (mainly Sarawak and Sabah), less than 1000 ha in Brunei, 5000 ha in the Philippines, 20 000 ha in Papua New Guinea, 5000 ha in Thailand and 10 000 ha on the Pacific islands. Most sago starch is consumed locally or traded on domestic markets. It accounts for less than 3% of international trade in starches. Some of it is traded as sago pearls: partially gelatinized kernels, 1—2 mm in diameter, obtained by forcing raw starch paste through a sieve and stirring the extruded pieces of paste on a hot-plate until hard and rounded. Sometimes, pearled starches of non-palm origin are erroneously called 'sago pearls' or even just 'sago'.
Unambiguous economic statistics are scarce. In 1992, the export of dry sago starch from Sarawak was 45 700 t (value MY$ 31 million). On Bengkalis (Riau Islands), an old centre of sago starch production, 30 mills operated in 1980 with a total output of 6600 t/year of dry starch. In the 1930s, 30 000—40 000 t/year wet starch production from the east coast of Sumatra was exported to Singapore to be refined and re-exported. The once prosperous sago-starch trade through Singapore has declined steadily since the 1950s.
Unambiguous economic statistics are scarce. In 1992, the export of dry sago starch from Sarawak was 45 700 t (value MY$ 31 million). On Bengkalis (Riau Islands), an old centre of sago starch production, 30 mills operated in 1980 with a total output of 6600 t/year of dry starch. In the 1930s, 30 000—40 000 t/year wet starch production from the east coast of Sumatra was exported to Singapore to be refined and re-exported. The once prosperous sago-starch trade through Singapore has declined steadily since the 1950s.
Properties
Purified sago starch consists of 27% amylose and 73% amylopectin. Dry starch samples from various parts of Indonesia contained water 10—17%, protein 0.31%, fat 0.11—0.25%, carbohydrates 81—88%, fibre 1.35%, ash 0.15—0.28%. A sago-based diet should be complemented with other foodstuffs to provide essential proteins, minerals and vitamins.
In Sarawak, the dry matter content of the grated whole pith (sample of 6 palms) contained: N 0.15%, P 0.046%, K 0.45%, Ca 0.24%, Mg 0.09% and starch 54%.
Sago weevil larvae from Sarawak and Papua New Guinea, weighing 3—8 g, contained: water 65—75%, protein 3—7%, fat 10—30%.
In Sarawak, the dry matter content of the grated whole pith (sample of 6 palms) contained: N 0.15%, P 0.046%, K 0.45%, Ca 0.24%, Mg 0.09% and starch 54%.
Sago weevil larvae from Sarawak and Papua New Guinea, weighing 3—8 g, contained: water 65—75%, protein 3—7%, fat 10—30%.
Description
A medium to tall palm tree, flowering once only, andromonoecious, forming basal suckers. Roots spongy but with a tough central fibro-vascular strand, not extending to great depth (80—100 cm in peat soil); pneumatophores (air roots) present. Trunk 30—60 cm in diameter, 7—20(—25) m tall, lower part ringed with leaf scars, upper part covered with semi-persistent leaf sheaths; epidermis thin, very sclerenchymatous, surrounding the fibrous bark, 5—10 mm thick; under the bark an extremely hard layer of sclerenchymatous fibre bundles, up to 1 cm thick surrounds the parenchymatous pith. Leaves 18—24 in vigorous trunked palms, simply pinnate, 5—7 m long (sometimes up to twice as long); petiole very robust, widening at its base into a stem-clasping sheath; sheath and petiole unarmed or armed to various degrees with needle-like spines, up to 22 cm long, arranged in transverse combs; leaflets up to 200 per leaf, 50—160 cm 3—6(—9) cm, often with small spines along the margins and on the midrib and sometimes with an apical, filiform appendage, margins usually valvate and reflexed. Inflorescence apparently a terminal panicle, 3—5(—7.5) m high and wide; first-order branches (10—)15—30, straight to curving upward, morphologically constituting separate lateral inflorescences arranged spirally on the main stem in the axils of reduced leaves or bracts with a phyllotaxy of 5/13, rigidly and distichously branched to the third order; the flower-bearing third-order branches spadix-like, rust-coloured when young, darker and more red from densely packed bulging flower buds later; flowers in pairs arranged spirally, each pair consisting of one male and one hermaphrodite flower, but up to half of the buds, usually most of them male, may abort before they reach anthesis; bracts of the first to the third order smooth to spinulescent outside; flowers 3-merous with 6 stamens. Fruit a depressed-globose to obconical drupe, 3.0—5.0(—7.0) cm in diameter, covered with 18(—19) vertical rows of scales, rhomboid, pointing downwards, greenish-yellow, turning straw-coloured towards or after fruit fall; scale layer lined inside with a white spongy layer. Seed subglobose, about 3 cm in diameter, firmly embedded in shiny cream-coloured firm flesh which turns pinkish when exposed to air; testa dark brown; endosperm homogeneous, horseshoe-shaped in longitudinal section because of large chalazal cavity; seeds often fail to develop, resulting in fruits filled with the cream-coloured flesh only.
Image
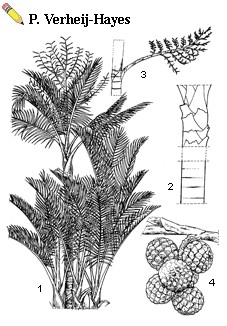 | Metroxylon sagu Rottboell - 1, habit of flowering palm with tillers; 2, stem [upper part covered with leaf bases, lower part with leaf scars]; 3, first-order inflorescence branch; 4, fruits |
Growth and Development
The seed is viable as soon as the fruit is shed but may quickly lose its viability through desiccation. In the field, germinating seeds are always fully ripe (brown testa and hard endosperm inside a straw-coloured husk), and are found on damp soil or even in a thin layer of water, under conditions of high relative humidity. Seeds usually germinate within 3 weeks. Vegetative growth is divided into a rosette stage of 3.5—6 years and a trunk stage of 4—14 years, depending on palm type and growing conditions. If these conditions are optimal, leaves form at a rate of 2 per month during the rosette stage, slowing down to 1 per month during the trunk stage; longevity of adult leaves is 18—24 months. Basal suckers are formed continuously, the first ones appearing already during the first year after germination. Starch is stored in the parenchyma (pith) of the trunk, which is gradually filled from the base upward. Maximum volumetric mass of the starch (kg per m3 of pith) has been found to be about 190 in Sarawak, 330 in the Sepik area of Papua New Guinea, and 280 on Seram in Indonesia. During flowering and especially during fruit development, the starch is translocated towards the inflorescence. After fruits have been shed, most of the starch has disappeared from the trunk.
The generative stage is heralded by the 'shooting' of the main stem, forming the main flowering axis: new internodes become longer, stem diameter and leaf size decrease and rate of leaf formation increases. The development of the inflorescence is phased: first the main axis develops, then the first-order branches, subsequently the second-order branches, etc. Anthesis of a male flower lasts 1 day, of a hermaphrodite flower 1—2 days; the anthesis period for the entire inflorescence lasts 50—60 days. Within one inflorescence, most male flowers have opened (and aborted the next day) before most hermaphrodite flowers open: sago palm is largely, but not strictly protandrous. Stamens in the male and hermaphrodite flowers look the same, but the viability of pollen in the hermaphrodite flowers is uncertain.
As germinating seeds are often found where two palms close to each other have flowered simultaneously, it has been assumed that sago palm is mainly a cross-pollinator.
Recent research findings suggest that sago palm is most probably largely self-incompatible, i.e. only palms which are genetically sufficiently different are well able to fertilize each other. This, in combination with the natural vegetative propagation method of the palm, the small chance of overlap of the anthesis periods of palms, and the condition that these palms should be growing at a distance small enough to allow for the insect-assisted pollen transfer, would account for the very low percentage of seeded fruits often encountered.
A yield of 5000 (seeded or unseeded) fruits is common in Sarawak. A sample-based estimate of the number of fruit in a palm on Ternate was 28 800.
It takes about 3 years from the outwardly visible start of the generative stage to the shedding of the fruits, after which the trunk dies. So, the total life span of a sago palm ranges between 11—23 years.
In the meantime, suckers of various ages, some already with a trunk, may have developed under the parent palm. Some suckers may form trunks up to several metres away from the parent palm, after first forming a prostrate stem.
The generative stage is heralded by the 'shooting' of the main stem, forming the main flowering axis: new internodes become longer, stem diameter and leaf size decrease and rate of leaf formation increases. The development of the inflorescence is phased: first the main axis develops, then the first-order branches, subsequently the second-order branches, etc. Anthesis of a male flower lasts 1 day, of a hermaphrodite flower 1—2 days; the anthesis period for the entire inflorescence lasts 50—60 days. Within one inflorescence, most male flowers have opened (and aborted the next day) before most hermaphrodite flowers open: sago palm is largely, but not strictly protandrous. Stamens in the male and hermaphrodite flowers look the same, but the viability of pollen in the hermaphrodite flowers is uncertain.
As germinating seeds are often found where two palms close to each other have flowered simultaneously, it has been assumed that sago palm is mainly a cross-pollinator.
Recent research findings suggest that sago palm is most probably largely self-incompatible, i.e. only palms which are genetically sufficiently different are well able to fertilize each other. This, in combination with the natural vegetative propagation method of the palm, the small chance of overlap of the anthesis periods of palms, and the condition that these palms should be growing at a distance small enough to allow for the insect-assisted pollen transfer, would account for the very low percentage of seeded fruits often encountered.
A yield of 5000 (seeded or unseeded) fruits is common in Sarawak. A sample-based estimate of the number of fruit in a palm on Ternate was 28 800.
It takes about 3 years from the outwardly visible start of the generative stage to the shedding of the fruits, after which the trunk dies. So, the total life span of a sago palm ranges between 11—23 years.
In the meantime, suckers of various ages, some already with a trunk, may have developed under the parent palm. Some suckers may form trunks up to several metres away from the parent palm, after first forming a prostrate stem.
Other Botanical Information
The distinction at species level of palms with and without spines (Metroxylon rumphii and Metroxylon squarrosum — spiny; Metroxylon sagu — spineless) cannot be upheld. The reduction of the number of taxa of lower order in true sago palm, however, from 21 to 4 in the latest revision of the genus, may be too drastic. For example, variation in life span, which appears to be mainly genetically determined, and which is one of the bases of the distinction of types by sago growers, is, unfortunately, not taken into account. Neither are other variations recognized, e.g. in leaf form, spine density, bark thickness, starch colour, and even starch taste.
The number of sago palm types distinguished by local sago growers increases eastwards: from 2 on Siberut island west of Sumatra and in Riau province, to 2—3 in Sarawak, to 4—5 on Ternate and Halmahera, to 5—8 in the Central Moluccas, to 10—13 on New Guinea island.
The number of sago palm types distinguished by local sago growers increases eastwards: from 2 on Siberut island west of Sumatra and in Riau province, to 2—3 in Sarawak, to 4—5 on Ternate and Halmahera, to 5—8 in the Central Moluccas, to 10—13 on New Guinea island.
Ecology
Sago palm is a tree of the per-humid tropical lowlands, occurring naturally up to 700 m above sea-level (up to 1200 m in Papua New Guinea). The best conditions for sago palm growth are an average temperature of at least 26°C, a relative humidity of 90% and an irradiance of about 9 MJ/m2 per day.
Natural stands of sago palm occur on swampy coastal plains, river floodplains and higher up on flat valley floors. When growing downstream along rivers, tidal influences may be part of the habitat of sago palms, and may affect the level and salinity of flood water or groundwater. Daily flooding is harmful to seedling growth, as is salinity corresponding to electric conductivities (EC) of over 1 S/m. (EC of sea water is 4.4 S/m). Occasional flooding, even with very saline water is tolerated, however. Although found on mineral, peat and muck soils, sago palm grows best on mineral soils with a high organic matter content (up to 30%).
In New Guinea, sago palms occur mainly in 4 vegetation types. Ranging from land inundated most of the year to less flood-prone lands, one may successively encounter sago palm — Phragmites swamp (groves of trunkless sago palms in dense stands of the reed Phragmites karka (Retz.) Trin. ex Steud.), sago palm swamp (dense stands of sago palms, most of them trunkless), and sago palm forest (sago palms in various stages of development mixed with dicotyledonous trees in various proportions). On peat soils that are dry most of the year, Campnosperma — sago palm forest (sago palms forming an understorey under a closed canopy of Campnosperma brevipetiolatum Volkens) can be found. The most numerous and largest trunks are found in the sago palm forest.
As the water becomes more brackish, sago palm often borders on stands of the more salinity-tolerant nipa palm (Nypa fruticans Wurmb).
Natural stands of sago palm occur on swampy coastal plains, river floodplains and higher up on flat valley floors. When growing downstream along rivers, tidal influences may be part of the habitat of sago palms, and may affect the level and salinity of flood water or groundwater. Daily flooding is harmful to seedling growth, as is salinity corresponding to electric conductivities (EC) of over 1 S/m. (EC of sea water is 4.4 S/m). Occasional flooding, even with very saline water is tolerated, however. Although found on mineral, peat and muck soils, sago palm grows best on mineral soils with a high organic matter content (up to 30%).
In New Guinea, sago palms occur mainly in 4 vegetation types. Ranging from land inundated most of the year to less flood-prone lands, one may successively encounter sago palm — Phragmites swamp (groves of trunkless sago palms in dense stands of the reed Phragmites karka (Retz.) Trin. ex Steud.), sago palm swamp (dense stands of sago palms, most of them trunkless), and sago palm forest (sago palms in various stages of development mixed with dicotyledonous trees in various proportions). On peat soils that are dry most of the year, Campnosperma — sago palm forest (sago palms forming an understorey under a closed canopy of Campnosperma brevipetiolatum Volkens) can be found. The most numerous and largest trunks are found in the sago palm forest.
As the water becomes more brackish, sago palm often borders on stands of the more salinity-tolerant nipa palm (Nypa fruticans Wurmb).
Propagation and planting
Sago palm is mostly propagated from suckers. Rooted suckers about 1 year old with a basal diameter of 8—15 cm are severed from selected parent palms with a clean vertical cut through the runner (rhizomatous stem), leaving some 15 cm of the runner attached to the sucker to serve as food reserve. The cut wound is sometimes rubbed with wood ash to prevent rot. Treating the wound with a broad-spectrum fungicide has been shown to increase viability of the sucker. The runner with the roots should be kept from drying out. Usually, all the leaves are cut off; sometimes the spear and all or part of the youngest unfurled leaf are left on the sucker. Before planting in the field, suckers can first be kept in nurseries, either in polythene bags, or by simply putting them in shallow water, or if the water is deeper by tying them to a raft with only the runner and roots hanging in the water. Usually, only about half the propagated suckers are successful. The sucker survival rate may be increased by reducing the time between cutting the suckers and putting them in a nursery, by treating them with pesticide to prevent Rhynchophorus attack, and by shading them in the dry season. Propagation from seed has a considerably higher rate of success but viable seeds are difficult to obtain and the heterogeneity of the offspring, e.g. spininess, is a drawback.
Suckers are planted in the field at 6 m 6 m to 7 m 7 m. Planting depth is critical: the runner and especially the cut end should be buried to prevent Rhynchophorus attack and desiccation, but the shoot and especially the apical meristem inside it should be above water table (or above flood-water level), also in the wet season, to prevent rot. If necessary, the suckers are staked. Plenty of shade should be provided.
In vitro propagation of sago palms is still in the experimental stage. Plantlets have been derived from embryo culture, but micro-propagation through multiple shoot induction from sago explants has not yet been successful.
Suckers are planted in the field at 6 m
In vitro propagation of sago palms is still in the experimental stage. Plantlets have been derived from embryo culture, but micro-propagation through multiple shoot induction from sago explants has not yet been successful.
Husbandry
The sucker is established as soon as the spear plus a new leaf have unfolded, normally within 3 months. Shade is then gradually removed.
Weeding is necessary until the leaf canopy has closed. Old leaves are pruned and used as mulch. One sucker is allowed to develop into a trunk every 2 years if clump spacing is 6 m 6 m, or every one and a half years if the spacing is 7 m 7 m. All other suckers are pruned. Thus, an annual yield of 136—139 trunks per hectare may be achieved. The water table should be no lower than 50 cm. Fertilizers are normally not used; in Peninsular Malaysia the palms grow on flood-prone river banks, the river water probably carries all the necessary nutrients. Deficiencies have been shown to reduce the rate of leaf formation and the leaf area of new leaves in seedlings.
Weeding is necessary until the leaf canopy has closed. Old leaves are pruned and used as mulch. One sucker is allowed to develop into a trunk every 2 years if clump spacing is 6 m
Diseases and Pests
In extensive exploitation from semi-wild stands, none of the pests encountered are economically important. In intensive estate cultivation in Sarawak, however, many pests have become economically important, the most important being the larvae of the hispid beetle Botryonopa grandis, which attack the soft tissues of furled leaves, termites (Coptotermes sp.), which tunnel through the young trunk up to the growing point, and the larvae of the red-striped palm weevil Rhynchophorus schach (= Rhynchophorus ferrugineus var. schach), the so-called sago worms, which eat away at the trunk's pith. Other pests reported from elsewhere include Rhynchophorus ferrugineus from Siberut (Indonesia), rhinoceros beetle (Oryctes centaurus) from Papua New Guinea, and nettle caterpillars (Darna spp.) from Java.
Harvesting
To get the most starch out of one palm, it should be harvested at the peak of its starch content, which is reached during the generative stage some time between the beginning of anthesis and the beginning of fruit development. To maximize starch production per unit time, however, trees should be felled before the inflorescence emerges, when starch accumulation rate has not yet slackened. After felling, the crown is severed from the trunk and the old leaf sheaths are removed. The leaf bearing part of the trunk contains little starch.
Yield
Top annual yield of dry starch from a first crop of palms of short life cycle in Peninsular Malaysia is about 25 t/ha, equivalent to 138 trunks of 180 kg each. Yields of the subsequent ratoon crops stabilize at about 15 t/ha (85 trunks of 180 kg each). Recorded production of dry starch from single 'mature' trunks in uncultivated stands range from 20—400(—800) kg. The production capacity of semi-wild stands is estimated at 50 trunks per ha per year, producing 10 t/ha, whereas good quality wild stands on the drier parts of swamps are estimated to produce 25 trunks per ha per year, yielding 5 t/ha.
Handling After Harvest
Processing consists of separation of bark and pith, pulverization of the pith and separation of the starch grains from the other pith constituents. Traditionally, most of the processing is done at the felling site. The trunk is split lengthwise with wedges or partly debarked (i.e. the bark proper plus the outer hard layer of fibre bundles is removed). The exposed pith is pounded loose and pulverized with a hoe-like or adze-like instrument or grated with a nail-studded plank. The starch grains are leached out of the pulverized pith with water over a sieve and the starch is recovered from the slurry passing through the sieve by letting it settle. Pith starts fermenting spontaneously soon after it is pulverized, giving off an acid smell and causing irreversible staining of the starch. So starch extraction should follow pith pulverization as soon as possible.
Traditionally, only the wet starch (starch content 60%) is removed from the field. In planted stands, the trunks are usually cut into lengths of about 1 m. These logs (starch content 20—25%), weighing 80—120 kg, are rolled and floated to a central mill for further processing, a network of waterways being indispensable.
Traditionally, only the wet starch (starch content 60%) is removed from the field. In planted stands, the trunks are usually cut into lengths of about 1 m. These logs (starch content 20—25%), weighing 80—120 kg, are rolled and floated to a central mill for further processing, a network of waterways being indispensable.
Genetic Resources
In Sarawak (Department of Agriculture, and Land Custody and Development Authority) and in the Moluccas, Indonesia (Makariki Experiment Station of the Coconut Research Institute at Seram Island), a start has been made with the collection of sago palm types from Eastern Indonesia and Papua New Guinea.
Breeding
For estate cultivation, palm types are needed which (1) have a short life cycle (brief rosette stage and quick 'maturation') to allow an early first harvest, (2) have a high starch accumulation rate (high yield per unit time), (3) can be planted densely (high yield per unit area), (4) are responsive to fertilizers, (5) have pest and disease resistance/tolerance, and (6), especially in Sarawak, are able to grow well on peat soil.
Prospects
Sago palm is one of the underexploited trees in South-East Asia. Sago starch is mainly harvested from wild or semi-wild (i.e. planted but neglected) stands. Vast areas of natural sago palm stands, in New Guinea in particular, are left unused because of the inaccessible habitat and their remoteness. Until the mid 1980s, sago palm had been cultivated regularly only in Peninsular Malaysia, especially in the State of Johor. Since about 1970, international interest in sago palm as a plant resource in equatorial swamps has increased and desk studies have demonstrated its economic viability as a plantation crop. In 1987, the development of an 7700 ha sago plantation was started near Mukah, Sarawak, on deep peat. A new plantation has also been opened up in Indonesia in Riau province. New large-scale schemes exploiting natural stands have come into operation in Indonesia on Halmahera and in Irian Jaya.
At present research is intensifying. In Sarawak, an experiment station devoted to research on growing sago palm on deep peat has been operating since 1982. Indonesia is planning to establish a sago palm research station.
At present research is intensifying. In Sarawak, an experiment station devoted to research on growing sago palm on deep peat has been operating since 1982. Indonesia is planning to establish a sago palm research station.
Literature
FAO, 1986. The development of the sago palm and its products. Report of the FAO/BPPT Consultation; Jakarta, Indonesia, 16-21 January 1984. Food and Agriculture Organization (FAO), Rome, Italy. 252 pp.
Flach, M., 1983. The sago palm. FAO Plant Production & Protection Paper No 47. Food and Agriculture Organization (FAO), Rome, Italy. 85 pp.
Flach, M. & Schuiling, D.L., 1989. Revival of an ancient starch crop: a review of the agronomy of the sago palm. Agroforestry Systems 7(3): 259-281.
Ng Thai Tsiung, Tie Yiu Liong & Kueh Hong Siong (Editors), 1991. Towards greater advancement of the sago industry in the '90s. Proceedings of the Fourth International Sago Symposium, 6-9 August, 1990, Kuching, Sarawak, Malaysia. Ministry of Agriculture and Community Development, Kuching, Sarawak, Malaysia. 225 pp.
Rauwerdink, J.B., 1986. An essay on Metroxylon, the sago palm. Principes 30(4): 165-180.
Schuiling, D.L., Jong, F.S. & Flach, M., 1993. Exploitation and natural variability of the sago palm (Metroxylon sagu Rottb.): Report of a Sarawak and all-Indonesia study tour, January-February 1992. Department of Agronomy, Wageningen Agricultural University, the Netherlands. 82 pp.
Stanton, W.R. & Flach, M. (Editors), 1980. Sago: the equatorial swamp as a natural resource. Proceedings of the Second International Sago Symposium, Kuala Lumpur, 1979. Nijhoff, the Hague, the Netherlands. 244 pp.
Tan, K. (Editor), 1977. Sago-76: Papers of the First International Sago Symposium 'The Equatorial Swamp as a Natural Resource'. Kuching (Malaysia), 1976. Kemajuan Kanji (now: Kemikro Bhd), Kuala Lumpur, Malaysia. 330 pp.
van Kraalingen, D.W.G., 1984. Some observations on sago palm growth in East Sepik River Basin (Papua New Guinea). DME/EPU Report No 1/84. Department of Minerals and Energy, Energy Planning Unit, Konedobu, Papua New Guinea. 69 pp.
Yamada, N. & Kainuma, K. (Editors), 1986. Sago-'85, the Third International Sago Symposium. Tokyo, Japan, May 20-23, 1985. The Sago Palm Research Fund. Tropical Agricultural Research Center, Tsukuba, Japan. 233 pp.
Flach, M., 1983. The sago palm. FAO Plant Production & Protection Paper No 47. Food and Agriculture Organization (FAO), Rome, Italy. 85 pp.
Flach, M. & Schuiling, D.L., 1989. Revival of an ancient starch crop: a review of the agronomy of the sago palm. Agroforestry Systems 7(3): 259-281.
Ng Thai Tsiung, Tie Yiu Liong & Kueh Hong Siong (Editors), 1991. Towards greater advancement of the sago industry in the '90s. Proceedings of the Fourth International Sago Symposium, 6-9 August, 1990, Kuching, Sarawak, Malaysia. Ministry of Agriculture and Community Development, Kuching, Sarawak, Malaysia. 225 pp.
Rauwerdink, J.B., 1986. An essay on Metroxylon, the sago palm. Principes 30(4): 165-180.
Schuiling, D.L., Jong, F.S. & Flach, M., 1993. Exploitation and natural variability of the sago palm (Metroxylon sagu Rottb.): Report of a Sarawak and all-Indonesia study tour, January-February 1992. Department of Agronomy, Wageningen Agricultural University, the Netherlands. 82 pp.
Stanton, W.R. & Flach, M. (Editors), 1980. Sago: the equatorial swamp as a natural resource. Proceedings of the Second International Sago Symposium, Kuala Lumpur, 1979. Nijhoff, the Hague, the Netherlands. 244 pp.
Tan, K. (Editor), 1977. Sago-76: Papers of the First International Sago Symposium 'The Equatorial Swamp as a Natural Resource'. Kuching (Malaysia), 1976. Kemajuan Kanji (now: Kemikro Bhd), Kuala Lumpur, Malaysia. 330 pp.
van Kraalingen, D.W.G., 1984. Some observations on sago palm growth in East Sepik River Basin (Papua New Guinea). DME/EPU Report No 1/84. Department of Minerals and Energy, Energy Planning Unit, Konedobu, Papua New Guinea. 69 pp.
Yamada, N. & Kainuma, K. (Editors), 1986. Sago-'85, the Third International Sago Symposium. Tokyo, Japan, May 20-23, 1985. The Sago Palm Research Fund. Tropical Agricultural Research Center, Tsukuba, Japan. 233 pp.
Author(s)
D.L. Schuiling & F.S. Jong
Correct Citation of this Article
Schuiling, D.L. & Jong, F.S., 1996. Metroxylon sagu Rottboell. In: Flach, M. & Rumawas, F. (Editors): Plant Resources of South-East Asia No 9: Plants yielding non-seed carbohydrates. PROSEA Foundation, Bogor, Indonesia. Database record: prota4u.org/prosea

All texts are licensed under a Creative Commons Attribution-Noncommercial-Share Alike 3.0 Netherlands License
This license does not include the illustrations (Maps,drawings,pictures); these remain all under copyright.